聴覚は、空気を媒質とした疎密波である音を電気信号である神経の活動信号に変換し、音の強弱・
音源の左右の定位・音色・音声言語等の様々な情報を認識する働きである。この機序には、大きく分けて、
伝音系と感音系がある。伝音系の働きは、外耳における共鳴効果による増幅効果と中耳におけるインピーダンス整合による
空気の疎密波から液体である内耳液の疎密波への変換の働きに分けられる。
感音系の働きは、内耳での液体の疎密波から神経の活動電位への変換と延髄から視床を経て大脳皮質に至るまでに行われる情報伝達の処理に分けられる。
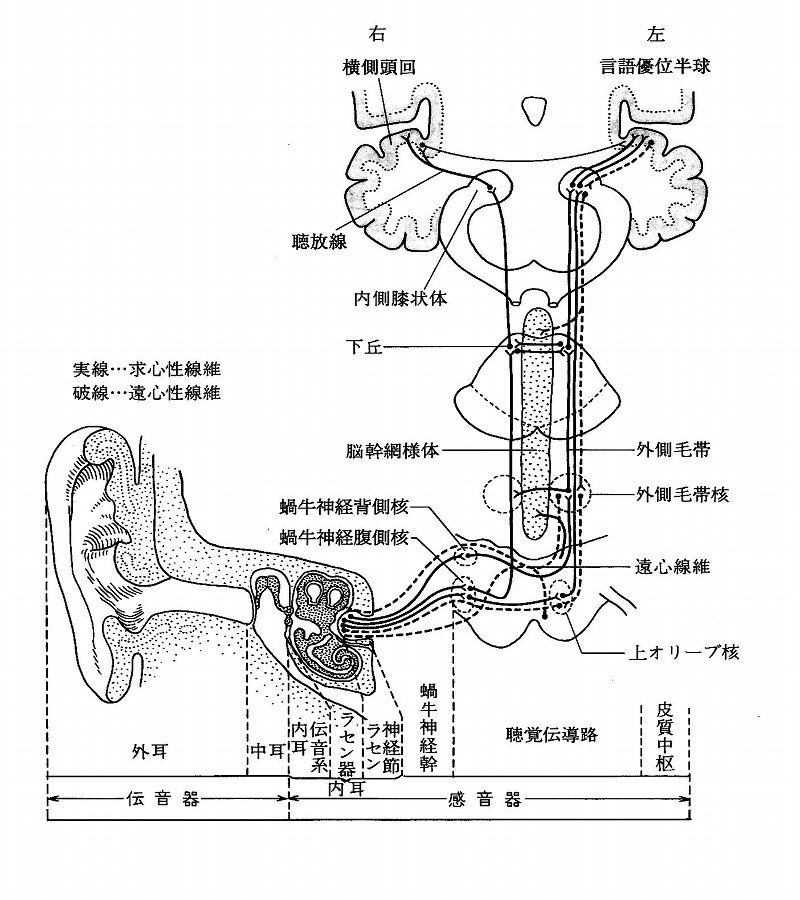
聴覚器は大きく外耳・中耳・内耳に分けられる。外耳は耳介軟骨に支えられた皮膚のヒダで、鼓膜に至るまでの外耳道を形成している。
中耳は主に鼓室・耳小骨からなる。鼓膜は、外耳道からつながる重層扁平上皮・線維束・中耳粘膜から連なる単層扁平上皮の3層からなる。
鼓室は耳小骨を入れた空洞となっており、内部は粘膜に覆われている。耳小骨は、ツチ骨・キヌタ骨・アブミ骨の3つの骨がそれぞれ関節で連合し、
耳小骨連鎖を形成している。また、ツチ骨前突起には前ツチ骨靱帯が、キヌタ骨短突起には後キヌタ骨靱帯が鼓室に固定されており、
耳小骨連鎖の振動の回転軸としての意味を持っている。内耳は、蝸牛・前庭・三半規管が、互いに交通し、内外リンパを満たした膜迷路を形成している。
外耳の共鳴効果は、一端が鼓膜で閉鎖された共鳴管としての働きによるもので、2.5kHz~4kHzの間に約10dB程度の増強効果がある。
この効果は、ヒトで見られる気導聴力の周波数毎の感度差の一因とされている。耳介自体には集音の働きはないが、両耳に到達する音の周波数
・位相が変化することで音源の立体的な定位に役立っている。
中耳でのインピーダンス整合は、空気から液体である内耳の外リンパに疎密波を変換する働きを持つ。
空気の疎密波が液体に達した場合、そのエネルギーの大部分は空気・液体の境界で反射されてしまい、液体に伝わるエネルギーはわずかとなってしまう。
この問題を解決するため、中耳では、ツチ骨、キヌタ骨、アブミ骨の耳小骨連鎖・耳小骨靱帯の形成するてこ比・面積比によって、
空気の振動を効率的に液体の振動に変換している。ツチ骨、キヌタ骨の間には、テコのような作用があり、長さにして1.3:1程度、
約2.5dBの増強効果がある。また、空気振動を受ける鼓膜の面積と液体振動を伝えるアブミ骨底の面積は、17:1程度、約25dBの増強効果がある。
以上、合計27.5dBの増強効果によって、中耳では、空気の疎密波が液体である内耳の外リンパの疎密波に効率的に変換されている。
内耳液での疎密波が神経の活動電位に変換されるのは、蝸牛基底板・有毛細胞の働きによる。
中耳より変換された内耳外リンパの疎密波は、基底板を振動させる。この振動は、基底板の特徴的な形状により、
周波数によって異なった基底板の部位に最大振幅を生じさせる。これは進行波と呼ばれ、高音では基底回転に近い部位で、
低音では蝸牛頂に近い部位で最大振幅が生じる。ここまでは内耳伝音系と呼ばれており、外有毛細胞や神経細胞による能動的な作用が無くても、
おおまかな周波数特性を持つ。
基底板上に生じた振幅は、基底板に接触している有毛細胞との間にずれ運動を引き起こし、有毛細胞の聴毛を屈曲する。
外有毛細胞の聴毛は蓋膜にささるように保持されており、基底板振動により直接ずれが生じる1)。内有毛細胞の聴毛は蓋膜とは接していないが、
この間の内リンパの流れにより聴毛が屈曲する。聴毛の屈曲により、聴毛のtip-linkの付け根に存在するイオンチャンネルが開き、
Kイオンが流入して脱分極が生じる2)3)。聴毛を最長側に屈曲させた時に細胞は最も刺激され、聴毛の周期的な屈曲により、
脱分極・過分極が繰り返される4)。この働きにより、機械的エネルギーが電気的エネルギーに変換される。外有毛細胞は、
膜電位の変化により、機械的に伸縮する働きがあり、基底板での振動をさらに増強し、周波数弁別能を高めている。
この外有毛細胞の働きは、基底板・前庭窓・耳小骨・鼓膜と通常とは逆向きに伝わり、耳からマイクロホンでとらえることができる(耳音響放射)。
蝸牛は、内リンパ・外リンパの二種類の細胞外液で満たされている。外リンパは通常の細胞外液とほぼ同じようなイオン組成であるが、
内リンパはカリウムイオンが約140mMと高濃度、ナトリウムイオンは約10mMと低濃度であり、カルシウムイオン濃度は非常に低くなっており、
外リンパと比べて約80mVと高電位になっている5)。基底板に存在する有毛細胞は、感覚毛を有する頂上の膜のみを内リンパに接しており、
音刺激の発生時に感覚毛に有するチャンネルがわずかに開くだけで大量のカリウムイオンが有毛細胞内に流入し、
刺激に対し敏感に反応することができるようになっている。また、このチャンネルは無音時でも10%程度開口しており、
音に対する感度が上がるようになっている。この内リンパ・外リンパ間のイオン濃度差・電位差は血管条によるカリウム・
ナトリウムの能動的な輸送とタイトジャンクションによる働きによるものである6)7)8)。
有毛細胞の直下では、蝸牛神経の樹状突起が神経終末となってシナプスを形成しており、有毛細胞からの神経伝達物質をスパイク放電に変換している。
音の入力により有毛細胞からの神経伝達物質が増加すると、スパイク放電の頻度が上昇し、高位ニューロンへと情報を伝達する。
各神経線維には、最も鋭敏に応じる周波数があり、特徴周波数と呼ばれている。この周波数の幅は、音圧によって異なる。
低音圧では、蝸牛の周波数特性と変わらないが、音圧が上がると、蝸牛神経から上位のニューロンになるに従って、
それぞれの神経線維の応じる周波数範囲は鋭くなり、内側膝状体で完成される。
蝸牛軸内に存在するらせん神経節から蝸牛神経背側核・腹側核、上オリーブ核、外側毛帯、下丘を経て内側膝状体に至るが、
この経路中で、左右での時間差・音圧差・周波数成分の差等の検出が行われている。哺乳類では、
内側上オリーブ核で左右の時間差情報が検出されており、
シナプスにより形成された遅延回路により両耳間の時間差が神経の発火頻度にコードされていると推定されている9)。
また、左右の音圧差は外側上オリーブ核で行われていると考えられ、同側の音で興奮し、
対側の音で抑制される細胞により音圧がコード化されると考えられる10)。周波数成分の差は、背側蝸牛神経核・下丘により行われている。
音源が移動すると、耳介によって周波数成分にノッチが生じ、下丘には特定の周波数帯が弱くなると特異的に興奮する細胞が存在するため、
音源の定位として知覚できる11)。以上の左右の時間差・音圧差・周波数成分の差等が下丘で統合され、
上丘では聴覚と視覚との情報が統合され空間地図が認知されていると考えられている。
下丘は、視床の神経核である内側膝状体に情報を入力するとともに、上位神経核から抑制性の入力を受けている。
内側膝状体でも、下位である下丘から周波数・音圧・両耳間の差についての情報を受け取ると共に、視床網様核・
大脳皮質と入出力の経路を持ち、ネットワークを形成している。
内側膝状体・視床網様核・大脳皮質が形成する回路は、内側膝状体での下丘からの情報を修飾し、時間軸に沿って情報を処理し、
大脳皮質に伝達する役割があると考えられる。内側膝状体は音反応に特化した神経核であると考えられてきたが、
近年、ラット等における実験から、その一部は体性感覚や視覚といった異種の感覚により影響を受けていることが分かってきた。
内側膝状体は、腹側亜核(MGBv)、内側亜核(MGBm)、背側亜核(MGBd)、膝上核(SG)に大きく分けられ、
それぞれにおいてニューロンの細胞膜特性や細胞形態に違いがある。MGBvには白色雑音刺激に対し、
短い潜時で反応するSL細胞と長い潜時で反応するLL細胞が認められており、SL細胞は下丘から音刺激に対する興奮性の刺激を受け取っているのに対し、
LL細胞は、音刺激により誘発される内側膝状体・視床網様核・大脳皮質のループの活動に応じて興奮が左右されると考えられている12)13)。
この働きにより、大脳皮質の高次の情報が内側膝状体で聴覚情報に重み付けされていると考えられる。
また、聴覚刺激への報酬による重み付けは、MGBdにおいても認められており、
MGBvでは体性感覚によって音刺激に対する反応が抑制される現象が確認されている14)。内側膝状体の何れの亜核においても、
下丘よりの音刺激の感覚入力と、先行して処理された感覚入力とがループ回路によって処理されており、
それらに加えて大脳で処理された高次の情報の入力や視覚・体性感覚といった異種の感覚入力が影響し、
周波数・音圧といった情報を処理し、一次聴覚野・二次聴覚野に伝達する働きを持っていると考えられる。
内側膝状体からは聴放線が形成され、同側の聴覚野に至っている。一次聴覚野は側頭葉の背側面にある横側頭回に存在し、
ブロードマンの41野にあたる。後方から前方に向けて、低音から高音に反応する領域が配列し、ここにおいても周波数局在性が認められている。
この一次聴覚野でも、視覚・触覚といった異種の感覚に応答する細胞による多感覚応答が見いだされており15)、
他の感覚との結合についても徐々に明らかになっている。
聴覚中枢の神経経路には、神経細胞の生理学的な特徴により分類されるClass 1とClass 2の2種類の経路がある。
Class 1経路は主要な聴覚情報を伝達する役割を持っていると考えられ、この経路のシナプスは、樹状突起は密で、軸索は太く、
神経終末は大きく、大きなEPSPによりイオンチャンネル型グルタミン酸受容体(iGluR)を興奮・抑制する。Class 2経路のシナプスは、
Class 1で伝達される情報を修飾する役割を持っていると考えられ、シナプスはClass 1に比べ、樹状突起は粗で、
軸索は細く、神経終末は小さく、EPSPは小さく、iGluRや代謝型グルタミン酸受容体(mGluR)を興奮・抑制する16)。
下丘から内側膝状体に至る経路では、下丘中心核(ICc)からMGBvに投射するClass 1の経路と、下丘背側核(ICd)・
下丘外側核(ICl)・下丘尾側核(ICca)からMGBd・MGBmに投射するClass 2との異なった経路が認められ、毛帯・非毛帯経路に分けられる16)17)。
内側膝状体から聴覚野への投射でも2種の経路が認められる。MGBv・MGBdから聴覚野の4層と6層に至る経路がClass 1経路にあたり、
主要な聴覚情報を伝達している。もう一つはMGBmから聴覚野の2層と3層に至る経路であり、聴覚情報を修飾するClass 2経路にあたる16)18)19)。
聴覚野から内側膝状体へ至る下行性の投射でも同様に2種の経路があると考えられる。聴覚野の5層からMGBdに至る経路では
Class 1経路の生理学的特徴が認められるのに対し、聴覚野の6層からMGBvに至る経路ではClass 2経路の特徴が認められる20)。
聴覚野の各層間や聴覚野間での経路の中にも同種の経路があると考えられており、徐々に明らかになってきている。
以上に述べたような働きにより、空気の疎密波が音の強弱・音源の左右の定位・音色・音声言語・空間等として認知される。 従来は聴覚のみに特化していると考えられてきた視床や聴覚野において、異種の感覚に対する応答が確認されるようになり、 視覚・体性感覚といった異種の感覚が統合され、聴覚が認識されていることも分かってきている。
(野村恭也:新耳鼻咽喉科学 改訂10版.p.27,南山堂,2004 より)
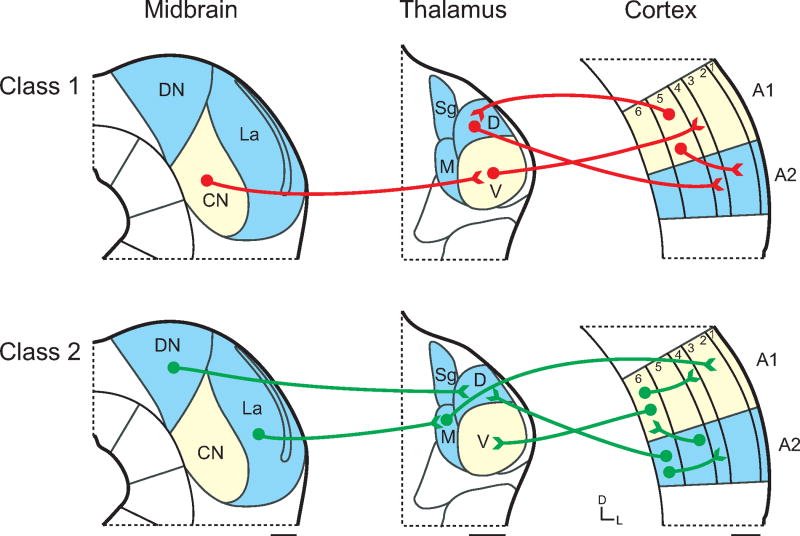
Class 1経路は主要な聴覚情報を伝達する役割を持っているのに対し、Class 2経路は主要な情報を修飾する役割を持っていると推定されている。
この2種類の経路は、下丘から内側膝状体・聴覚野に至る伝導路で確認されているが、その他の伝導路にも存在すると考えられている。
DN:下丘背側核、CN:下丘中心核、La:下丘外側核、Sg:内側膝状体膝上核、D:内側膝状体背側亜核、M:内側膝状体内側亜核、
V:内側膝状体腹側亜核、A1:一次聴覚野、A2:二次聴覚野
(Lee CC ら16)より)
1) Furness DN, Hackney CM. Cross-links between stereocilia in the guinea pig cochlea. Hear Res. 1985;18:177-88.
2) Glowatzki E, Fuchs PA. Transmitter release at the hair cell ribbon synapse. Nat Neurosci. 2002;5:147-54.
3) Gillespie PG, Müller U. Mechanotransduction by Hair Cells: Models, Molecules, and Mechanisms. Cell. 2009;139:33-44.
4) Kitajiri S, Sakamoto T, Belyantseva IA, et al. Actin-bundling protein TRIOBP forms resilient rootlets of hair cell stereocilia essential for hearing. Cell. 2010;141:786-98.
5) Georg v. Békésy. DC Resting Potentials Inside the Cochlear Partition. J Acoust Soc Am. 1952;24:72-6.
6) Salt AN, Melichar I, Thalmann R. Mechanisms of endocochlear potential generation by stria vascularis. Laryngoscope. 1987;97:984-91.
7) Ikeda K, Morizono T. Electrochemical profiles for monovalent ions in the stria vascularis: cellular model of ion transport mechanisms. Hear Res. 1989;39:279-86.
8) Nin F, Hibino H, Doi K, et al. The endocochlear potential depends on two K+ diffusion potentials and an electrical barrier in the stria vascularis of the inner ear. Proc Natl Acad Sci USA. 2008;105:1751-6.
9) Jeffress L A. A place theory of sound localization. J Comp Physiol Psychol. 1948;41:35-9.
10) Mogdans J, Knudsen EI. Representation of interaural level difference in the VLVp, the first site of binaural comparison in the barn owl's auditory system. Hear Res. 1994;74:148-64.
11) Nelken I, Young ED. Two separate inhibitory mechanisms shape the responses of dorsal cochlear nucleus type IV units to narrowband and wideband stimuli. J Neurophysiol. 1994;71:2446-62.
12) Huguenard JR, Prince DA. Intrathalamic rhythmicity studied in vitro: nominal T-current modulation causes robust antioscillatory effects. J Neurosci. 1994;14:5485-502.
13) Smith PH, Bartlett EL, Kowalkowski A. Unique combination of anatomy and physiology in cells of the rat paralaminar thalamic nuclei adjacent to the medial geniculate body. J Comp Neurol. 2006;496:314-34.
14) Donishi T, Kimura A, Imbe H, et al. Sub-threshold cross-modal sensory interaction in the thalamus: lemniscal auditory response in the medial geniculate nucleus is modulated by somatosensory stimulation. Neuroscience. 2011;174:200-15.
15) Lakatos P, Chen CM, O'Connell MN, et al. Neuronal oscillations and multisensory interaction in primary auditory cortex. Neuron. 2007;53:279-92.
16) Lee CC, Sherman SM. On the classification of pathways in the auditory midbrain, thalamus, and cortex. Hear Res. 2011;276:79-87.
17) Bartlett EL, Smith PH. Effects of paired-pulse and repetitive stimulation on neurons in the rat medial geniculate body. Neuroscience. 2002;113:957-74.
18) Huang CL, Winer JA. Auditory thalamocortical projections in the cat: laminar and areal patterns of input. J Comp Neurol. 2000;427:302-31.
19) Viaene AN, Petrof I, Sherman SM. Synaptic properties of thalamic input to layers 2/3 and 4 of primary somatosensory and auditory cortices. J Neurophysiol. 2011;105:279-92.
20) Winer JA, Diehl JJ, Larue DT. Projections of auditory cortex to the medial geniculate body of the cat. J Comp Neurol. 2001;430:27-55.
Recently, Richard Stallman gave a speech in which he illustrated an academic
point about programming history by quoting a guy who described vi as 'an
editor spread at sword-point and which is really hard to use'.
I think I speak for all moderate vi(m) users when I say -- DEATH and DAMNATION
(in that order) to this Cardinal of the CTRL key! Needless to say my own local
vim user group has dispatched assassins to kill Mr. Stallman, but this is
hardly the end of the story. The fact is that a man has referred to another
man who in turn expressed some often-voiced reservations about OUR EDITOR! On
behalf of all editors of text everywhere, I implore EMACS users to return to
the true path, lest you be burned at the stake and then go to hell, the Buffer
From Which There Is No Unloading. We'll see how productive you are then, with
your ctrl-meta-alt and your ELISP and your 'ring buffer', whatever THAT is.
Peace and love to all.
^C
^X
quit
q
QUIT
exit :exit
zz
ZZ
kahei on
[Slashdot](http://linux.slashdot.org/comments.pl?sid=196931&cid=16136657)
-- kahei
-- Slashdot Comment ( http://linux.slashdot.org/comments.pl?sid=196931&cid=16136657 )
<f00li5h> rindolf: are you coming out this way on tour some time?
<rindolf> f00li5h: to .au?
<f00li5h> yes!
<rindolf> f00li5h: don't think so.
<f00li5h> you can pay some of my rent for a bit ^_^
<rindolf> f00li5h: I'm out of job too.
<f00li5h> perfect timing!
<f00li5h> no commitments
* f00li5h is very good at constructing circular arguments due to
his skill in constructing circular arguments
-- Will rindolf come to Australia?
-- #perl, Freenode